Cariotipo
normal y de individuos con alteraciones cromosómicas.
Introducción
El cariotipo es el complemento cromosómico particular de un individuo y viene
definido por el número y morfología de los cromosomas en la metafase mitótica.
En la especie humana la dotación cromosómica es de 2n = 46 (22 pares de
autosomas y un par de cromosomas sexuales).
En
el cariotipo humano los cromosomas se ordenan de mayor a menor. Hay cromosomas
grandes, medianos y pequeños. Al ordenar los comosomas se constituyen 7 grupos
atendiendo no sólo al tamaño sino también a la forma de las parejas
cromosómicas, dentro del cariotipo humano podemos encontrar cromosomas
metacéntricos (tienen los dos brazos aproximadamente iguales en longitud),
submetacéntricos (con un brazo más pequeño que otro) y acrocéntricos (con un
brazo corto muy pequeño)
Utilizando
técnicas de tinción estándar los cromosomas aparecen uniformemente teñidos en
metafase y se clasifican en 7 grupos de la A a la G atendiendo a su longitud
relativa y a la posición del centrómero que define su morfología.
Los
autosomas se numeran del 1 al 22 ordenados por tamaños decrecientes y por la
posición del centrómero. Los cromosomas sexuales X e Y constituyen un par
aparte, independientemente del resto (por su tamaño, el cromosoma X se incluiría
en el grupo C, y el Y, en el grupo G). De esta forma el cariotipo humano queda
constituido así:
GrupoPares
cromosómicosCaracterísticas
A
1, 2 y 3 Cr. muy grandes casi metacéntricos (1 y 3
metacéntricos, pero 2 submetacéntrico)
B
4 y 5 Cr. grandes y submetacéntricos, con dos
brazos muy diferentes en tamaño
C
6, 7, 8, 9, 10, 11 y 12 Cr. medianos submetacéntricos
D
13, 14 y 15 Cr. medianos acrocéntricos con satélites
E
16, 17 y 18 Cr. pequeños, metacéntrico el 16 y
submetacéntricos 17, 18
F
19 y 20 Cr. pequeños y metacéntricos
G
21 y 22 Cr. pequeños y acrocéntricos, con
satélites.
X,
Y El cr. X es parecido al
6. El Y, al grupo G, pero sin satélites.
(Todos
los cromosomas autosómicos están ordenados en orden decreciente de tamaño,
excepto el cromosoma 21 que ahora se sabe que es más pequeño que el 22)
Sin
embargo, atendiendo solamente a estos parámetros no es posible identificar
inequívocamente cada par de cromosomas. Para ello es necesario utilizar
diferentes técnicas de bandeo cromosómico. Los distintos patrones de bandas que
se consiguen son constantes y específicos de cada técnica y determinan la
distribución de regiones cromosómicas que se revelan positiva o negativamente
según el método utilizado.
En
1956, Tijo y Levan determinaran el complemento cromosómico diploide del hombre
(2n = 46). En 1959 Lejeune describió la primera cromosomopatía o
enfermedad originada por una alteración cromosómica, el síndrome de Down
producido por una trisomía del cromosoma 21. Desde entonces, la Citogenética
Humana ha ido desarrollándose como una ciencia médica.
Hay
dos tipos fundamentales de cromosomopatías:
-
Variaciones cromosómicas estructurales: afectan a la estructura del cromosoma en
cuanto a la ordenación lineal de los genes. Aquí se incluyen deleciones,
duplicaciones, inversiones y translocaciones.
-
Variaciones cromosómicas numéricas: afectan al número de cromosomas. Incluyen
las poliploidías (triploidía: 3n; tetraploidía: 4n) y los diversos tipos de
aneuploidía (trisomías: 2n+1; monosomías: 2n-1).
Por
otra parte, las anomalías cromosómicas pueden afectar a los autosomas o a los
cromosomas sexuales.
Las
alteraciones cromosómicas más frecuentes en humanos son:
Anomalías
autosómicas:
Síndrome
de Down, por trisomía del cromosoma 21, translocación 21/21 o
translocación 14/21.
Síndrome
de Patau, por trisomía del par 13.
Síndrome
de Edwards, por trisomía del par 18.
Síndrome
Cri du Chat, por deleción parcial del brazo corto del cromosoma 5
(5p).
Síndrome
de DiGeorge, por deleción parcial del brazo largo del cromosoma 22
(22q11).
Cromosoma
Filadelfia, formado por una translocación entre los cromosomas 9 y
22.
Anomalías
de los cromosomas sexuales:
Síndrome
de Klinefelter, por una constitución XXY, XXXY, XXXXY.
Síndrome
XYY, cromosoma Y extra en varones.
Síndrome
de Turner, constitución X0.
Síndrome
XXX, cromosoma X extra en mujeres.
Actualmente se ha llegado a profundizar bastante en el conocimiento del
cariotipo humano y se sabe que es relativamente frecuente la aparición de
anomalías cromosómicas. Por ejemplo, cerca de un 25% de los abortos ocurridos
antes de la octava semana de gestación tienen cariotipos anormales y un 0,5% de
los recién nacidos presentan aneuploidías.
Estas alteraciones no sólo pueden producir anomalías en el propio individuo
portador sino que, por tratarse de anomalías genéticas, pueden transmitirse a la
descendencia en el caso de que afecten a las células germinales. La detección
anticipada de anomalías cromosómicas permite dictaminar las posibilidades de que
la descendencia de una pareja portadora de una de ellas pueda presentarla o no.
Para ello es preciso conocer el cariotipo de cada progenitor, lo que permite
emitir un diagnóstico de su posible descendencia, con lo que el individuo será
consciente de sus posibilidades.
El
estudio del cariotipo tiene también su aplicación en el diagnóstico prenatal. Es
posible determinar la constitución cromosómica del feto antes de su nacimiento
pudiendo así observarse si presenta alguna anomalía cromosómica detectable. Hoy
en día, el diagnóstico prenatal se practica a
posteriori del inicio de la gestación y los resultados positivos suelen
plantear conflictos éticos y emocionales.
Se conoce como síntesis de proteínas al proceso por el cual se componen nuevas proteínas a partir de los veinte aminoácidos esenciales. En estre proceso, se transcribe el ADN en ARN. La síntesis de proteínas se realiza en los ribosomas situados en el citoplasma celular.
En el proceso de síntesis, los aminoácidos son transportados por ARN de transferencia correspondiente para cada aminoácido hasta el ARN mensajero donde se unen en la posición adecuada para formar las nuevas proteínas.
Al finalizar la síntesis de una proteína, se libera el ARN mensajero y puede volver a ser leido, incluso antes de que la síntesis de una proteína termine, ya puede comenzar la siguiente, por lo cual, el mismo ARN mensajero puede utilizarse por varios ribosomas al mismo tiempo.
A continuación puedes ver más información sobre en qué consiste el proceso de la síntesis de proteínas, cuales son sus fases y los pasos que se realizan en cada fase de la síntesis de proteínas.
Fases de las síntesis de proteínas
La realización de la biosíntesis de las proteínas, se divide en las siguientes fases:
Fase de activación de los aminoácidos.
Fase de traducción que comprende:
Inicio de la síntesis proteica.
Elongación de la cadena polipeptídica.
Finalización de la síntesis de proteínas.
Asociación de cadenas polipeptídicas y, en algunos casos, grupos prostésicos para la constitución de las proteínas.
Fase de activación de los aminoácidos
Mediante la enzima aminoacil-ARNt-sintetasa y de ATP, los aminoácidos pueden unirse ARN específico de transferencia, dando lugar a un aminoacil-ARNt. En este proceso se libera AMP y fosfato y tras él, se libera la enzima, que vuelve a actuar.
Inicio de la síntesis proteica
En esta primera etapa de síntesis de proteínas, el ARN se une a la subunidad menor de los ribosomas, a los que se asocia el aminoacil-ARNt. A este grupo, se une la subunidad ribosómica mayor, con lo que se forma el complejo activo o ribosomal.
Leer más sobre la fase de iniciación de la síntesis de proteínas.
Elongación de la cadena polipeptídica
El complejo ribosomal tiene dos centros o puntos de unión. El centro P o centro peptidil y el centro A. El radical amino del aminoácido inciado y el radical carboxilo anterior se unen mediante un enlace peptídico y se cataliza esta unión mediante la enzima peptidil-transferasa.
De esta forma, el centro P se ocupa por un ARNt carente de aminoácido. Seguidamente se libera el ARNt del ribosoma produciéndose la translocación ribosomal y quedando el dipeptil-ARNt en el centro P.
Al finalizar el tercer codón, el tercer aminoacil-ARNt se sitúa en el centro A. A continuación se forma el tripéptido A y después el ribosoma procede a su segunda translocación. Este proceso puede repetirse muchas veces y depende del número de aminoácidos que intervienen en la síntesis.
Leer más sobre la fase de elongación de la síntesis de proteínas.
Finalización de la síntesis de proteínas.
En la finalización de la síntesis de proteínas, aparecen los llamados tripletes sin sentido, también conocidos como codones stop. Estos tripletes son tres: UGA, UAG y UAA. No existe ARNt tal que su anticodón sea complementario. Por ello, la síntesis se interrumpe y esto indica que la cadena polipeptídica ha finalizado.
Leer más sobre la fase de terminación de la síntesis de proteínas.
La transcripción del ADN es el primer proceso de la expresión génica, mediante el cuál se transfiere la información contenida en la secuencia del ADN hacia la secuencia de proteína utilizando diversos ARN como intermediarios. Durante la transcripción genética, las secuencias de ADN son copiadas a ARN mediante una enzima llamada ARN polimerasa que sintetiza un ARN mensajero que mantiene la información de la secuencia del ADN. De esta manera, la transcripción del ADN también podría llamarse síntesis del ARN mensajero. Transcripción en eucariotas
En el caso de las eucariotas, el proceso se realiza en el núcleo, y es similar al de las procariotas, pero de mayor complejidad. Diferentes ARNp transcriben distintos tipos de genes. La ARNpII transcribe los pre-ARNm, mientras que la ARNpI y ARNpIII transcriben los ARN-ribosomales y ARNt, respectivamente. Los ARNs transcritos son modificados posteriormente. El pre-ARNm,por ejemplo, sufre un proceso de maduración que tras cortes y empalmes sucesivos elimina ciertos segmentos del ADN llamados los intrones para producir el ARNm final. Durante este proceso de maduración se puede dar lugar a diferentes moléculas de ARN, en función de diversos reguladores. Así pues, un mismo gen o secuencia de ADN, puede dar lugar a diferentes moléculas de ARNm y por tanto, producir diferentes proteínas. Otro factor de regulación propio de las células eucariotas son los conocidos potenciadores (en inglés: "enhancers"), que incrementan mucho (100 veces) la actividad de transcripción de un gen, y no depende de la ubicación de éstos en el gen, ni la dirección de la lectura.
Etapas de la transcripción
Clásicamente se divide el proceso de la transcripción en 3 etapas principales (iniciación, elongación y terminación), pero realmente se pueden diferenciar 5 etapas :
Preiniciación
Al contrario de la replicación de ADN, durante el inicio de la transcripción no se requiere la presencia de un cebador para sintetizar la nueva cadena, de ARN en este caso. Antes del inicio de la transcripción se necesita toda una serie de factores de transcripción que ejercen los factores de iniciación. Estos se unen a secuencias específicas de ADN para reconocer el sitio donde la transcripción ha de comenzar y se sintetice el ARN cebador. Esta secuencia de ADN en la que se ensamblan los complejos de transcripción se llama promotor. Los promotores se localizan en los extremos 5'-terminales de los genes, antes del comienzo del gen, y a ellos se unen los factores de transcripción mediante fuerzas de Van der Waals y enlaces de hidrógeno. Los promotores tienen secuencias reguladoras definidas, muy conservadas en cada especie, donde las más conocidas son la caja TATA (situada sobre la región -10), con la secuencia consenso TATA(A/T)A(A/T); y la caja TTGACA (situada en el punto -35). La formación del complejo de transcripción se realiza sobre el promotor TATA, allí se forma el núcleo del complejo de iniciación. Sobre la caja TATA se fija una proteína de unión (TBP) junto con el factor de transcripción TFII D (TF proviene del inglés: transcription factor). Después, a ellos se unen otros factores de transcripción específicos: TFII B se une a TBP, TFII A (opcional), que estabiliza el complejo TFII B-TBP; luego se une el complejo TFII F y ARN polimerasa, y al final TFII E y TFII H. Todo ello forma un complejo que se llama complejo de preiniciación cerrado o PIC. Cuando la estructura se abre por mediación del factor de transcripción TFII H, da comienzo la iniciación y al complejo abierto (por su acción helicasa dependiente de ATP).
Iniciación
Primero, una Helicasa separa las hebras de ADN en estas denominadas cajas TATA, ya que entre adenina y timina se establecen dos enlaces de hidrógeno, mientras que entre citosina y guanina se forman tres. Posteriormente se unen los factores y las proteinas de transcripción (TBP, TF2D, TF2B) permitiendo, de esta manera, el acceso de la ARN polimerasa al molde de ADN de cadena simple, siendo esta la ultima en posicionarse. Aunque la búsqueda del promotor por la ARN polimerasa es muy rápida, la formación de la burbuja de transcripción o apertura del ADN y la síntesis del cebador es muy lenta. La burbuja de transcripción es una apertura de ADN desnaturalizado de 18 pares de bases, donde empieza a sintetizarse el ARN cebador a partir del nucleótido número 10 del ADN molde de la burbuja de transcripción. La burbuja de transcripción se llama complejo abierto. La ARN polimerasa es una enzima formada por 5 subunidades: 2 subunidades α, 1 subunidad β, 1 subunidad β' y 1 subunidad ω que tiene como función la unión de ribonucleótidos trifosfato. Cuando se forma el complejo abierto, la ARN polimerasa comienza a unir ribonucleótidos mediante enlaces fosfodiéster, y una vez que se forma el primer enlace fosfodiéster, acaba la etapa de iniciación.y comienza asi la siguiente etapa.
Disgregación del promotor
Una vez sintetizado el primer enlace fosfodiéster, se debe deshacer el complejo del promotor para que quede limpio para volver a funcionar de nuevo. Durante esta fase hay una tendencia a desprenderse el transcrito inicial de ARN y producir transcritos truncados, dando lugar a una iniciación abortada, común tanto en procariontes como eucariontes. Una vez que la cadena transcrita alcanza una longitud de unos 23 nucleótidos, el complejo ya no se desliza y da lugar a la siguiente fase, la elongación.
La disgregación del promotor coincide con una fosforilación de la serina 5 del dominio carboxilo terminal de la ARN polimerasa, que es fosforilado por el TFII H (que es una proteína quinasa dependiente de ATP)
Elongación
La ARN polimerasa cataliza la elongación de cadena del ARN. Una cadena de ARN se une por apareamiento de bases a la cadena de ADN, y para que se formen correctamente los enlaces de hidrógeno que determina el siguiente nucleótido del molde de ADN, el centro activo de la ARN polimerasa reconoce a los ribonucleótidos trifosfato entrantes. Cuando el nucleótido entrante forma los enlaces de hidrógeno idóneos, entonces la ARN polimerasa cataliza la formación del enlace fosfodiéster que corresponde. A esto se le llama elongación, la segunda etapa de la transcripción del ARN.
Terminación
Al finalizar la síntesis de ARNm, esta molécula ya se ha separado completamente del ADN (que recupera su forma original) y también de la ARN polimerasa, terminando la transcripción. La terminación es otra etapa distinta de la transcripción, porque justo cuando el complejo de transcripción se ha ensamblado activamente debe desensamblarse una vez que la elongación se ha completado. La terminación está señalizada por la información contenida en sitios de la secuencia del ADN que se está transcribiendo, por lo que la ARN polimerasa se detiene al transcribir algunas secuencias especiales del ADN. Estas secuencias son ricas en guanina y citosina, situadas en el extremo de los genes, seguidas de secuencias ricas en timina, formando secuencias palindrómicas, que cuando se transcriben el ARN recién sintetizado adopta una estructura en horquilla que desestabiliza el complejo ARN-ADN, obligando a separarse de la ARN polimerasa, renaturalizándose la burbuja de transcripción. Algunas secuencias de ADN carecen de la secuencia de terminación, sino que poseen otra secuencia a la que se unen una serie de proteínas reguladoras específicas de la terminación de la transcripción como rho.
El proceso de replicación de ADN es el mecanismo que permite al ADN duplicarse (es decir, sintetizar una copia idéntica). De esta manera de una molécula de ADN única, se obtienen dos o más "clones" de la primera. Esta duplicación del material genético se produce de acuerdo con un mecanismo semiconservativo, lo que indica que las dos cadenas complementarias del ADN original, al separarse, sirven de molde cada una para la síntesis de una nueva cadena complementaria de la cadena molde, de forma que cada nueva doble hélice contiene una de las cadenas del ADN original. Gracias a la complementación entre las bases que forman la secuencia de cada una de las cadenas, el ADN tiene la importante propiedad de reproducirse idénticamente, lo que permite que la información genética se transmita de una célula madre a las células hijas y es la base de la herencia del material genético.
La molécula de ADN se abre como una cremallera por ruptura de los puentes de hidrógeno entre las bases complementarias liberándose dos hebras y la ADN polimerasa sintetiza la mitad complementaria añadiendo nucleótidos que se encuentran dispersos en el núcleo. De esta forma, cada nueva molécula es idéntica a la molécula de ADN inicial.
La replicación empieza en puntos determinados: los orígenes de replicación. Las proteínas iniciadoras reconocen secuencias de nucleótidos específicas en esos puntos y facilitan la fijación de otras proteínas que permitirán la separación de las dos hebras de ADN formándose una horquilla de replicación. Un gran número de enzimas y proteínas intervienen en el mecanismo molecular de la replicación, formando el llamado complejo de replicación o replisoma. Estas proteínas y enzimas son homólogas en eucariotas y arqueas, pero difieren en bacterias.
Características generales
Semiconservadora
En cada una de las moléculas hijas se conserva una de las cadenas originales, y por eso se dice que la replicación del ADN es semiconservadora. Hasta que finalmente se pudo demostrar que la replicación es semiconservadora, se consideraron tres posibles modelos para el mecanismo de la replicación:
Semiconservadora (modelo correcto). En cada una de las moléculas hijas se conserva una de las cadenas originales.
Conservadora. Se sintetiza una molécula totalmente nueva, copia de la original.
Dispersora, o dispersante. Las cadenas hijas constan de fragmentos de la cadena antigua y fragmentos de la nueva.
El experimento de Meselson y Stahl en 1958 permitió demostrar que el mecanismo real se ajusta a la hipótesis de replicación semiconservadora. Para ello se hicieron crecer células de Escherichia coli en presencia de nitrógeno-15, un isótopo del nitrógeno más pesado de lo habitual. En consecuencia, el isótopo se incorporó a las cadenas de ADN que se iban sintetizando, haciéndolas más pesadas.
Una vez conseguido el primer objetivo, las células fueron transferidas a un medio que contenía nitrógeno-14, es decir, un medio más ligero, donde continuaron su crecimiento (división celular, que requiere la replicación del ADN). Se purificó el ADN y se analizó mediante una centrifugación en gradiente de cloruro de cesio, en donde hay más densidad en el fondo del tubo que en la parte media del mismo.
En la primera generación se obtuvo una única banda de ADN con densidad intermedia. En la segunda generación se obtuvieron dos bandas, una con densidad ligera y otra con densidad intermedia o híbrida. En la tercera generación se obtuvieron dos bandas, una ligera (con una abundancia del 75%) y otra intermedia (con el 25% restante).
La banda intermedia o híbrida representa una molécula de ADN que contiene una cadena pesada (original) y otra ligera (recién sintetizada). Las cadenas ligeras representan una molécula de ADN en la que las dos cadenas han sido sintetizadas (no existían aun cuando las células se pusieron en presencia de nitrógeno-15.
El hecho de que cada vez haya más moléculas ligeras y se mantenga el número de moléculas intermedias demuestra que la replicación del ADN es semiconservadora. Si fuera conservadora, aparecería siempre una banda pesada y el resto ligeras (figuras 1.a, 1.b, 1.c) . Si fuera dispersante sólo aparecerían bandas híbridas de densidad intermedia en todas las generaciones.
Secuencial y bidireccional desde puntos fijos.
Los orígenes de replicación son los puntos fijos a partir de los cuales se lleva cabo la replicación, que avanza de forma secuencial formando estructuras con forma de horquilla. Por otro lado, la replicación se lleva a cabo bidireccionalmente, es decir, a partir de cada origen se sintetizan las dos cadenas en ambos sentidos.
El origen de replicación
La cantidad de ADN que se puede sintetizar a partir de un único origen de replicación se denomina replicón o unidad funcional de replicación. El genoma bacteriano es un replicón único circular. En organismos eucarióticos, la replicación del ADN se inicia en múltiples orígenes a la vez (hay uno cada 20 kb aproximadamente), es decir, hay varios replicones.
Los experimentos realizados por Cairns (1963) con bacterias Escherichia coli permitieron determinar la existencia de ese punto fijo u origen de replicación a partir del cual el genoma empezaba a replicarse. Los experimentos consistían en mantener un cultivo de E. coli creciendo en un medio que contenía timidina tritiada (timina marcada con tritio), de forma que el ADN quedara marcado radiactivamente pudiendo efectuarse una autorradiografía. A continuación se observaba al microscopio. Los resultados indicaban que la replicación en E. coli se iniciaba en un punto concreto (OriC).
Secuencialidad
Sueoka y Yoshikawa (1963) realizaron estudios genéticos de complementación de mutaciones que permitieron determinar que desde los orígenes la replicación avanza de forma secuencial. Trabajaron con Bacillus subtilis porque era posible obtener cultivos sincronizados de forma que todas las células del cultivo estuvieran en la misma fase del ciclo celular. El método consistía en la conjugación bacteriana de cepas silvestres con cepas mutantes incapaces de sintetizar determinados aminoácidos. Conociendo la localización de los genes que codifican las proteínas implicadas en la síntesis de los diferentes aminoácidos en el cromosoma bacteriano, y haciendo crecer las bacterias receptoras en un "medio mínimo" (donde sólo pudiesen crecer las que hubieran recibido alguno de estos genes), al extraer ADN a diferentes tiempos se observó que, tras la última extracción aparecía con mayor frecuencia el gen implicado en la síntesis de uno de los aminoácidos (el correspondiente a la "posición 1"), que el gen adyacente implicado en la síntesis de otro aminoácido ("posición 2"). De la misma forma, el gen que ocupaba la "posición 3" aparecía con menor frecuencia que el que ocupaba la "posición 2", y así sucesivamente.
Como los primeros genes en replicarse en la bacteria donadora serían los primeros en transferirse, el experimento permitió demostrar, a partir de las frecuencias relativas de los diferentes genes en las bacterias receptoras, que la replicación sigue un orden (es secuencial).
La replicación avanza en forma de horquilla
Debido a que en la célula ambas cadenas de la doble hélice de ADN se duplican al mismo tiempo, éstas deben separarse para que cada una de ellas sirva de molde para la síntesis de una nueva cadena. Por eso, la replicación avanza con una estructura en forma de horquilla formándose una burbuja u ojo de replicación (también llamada estructura θ cuando el ADN es circular debido a la similaridad entre la letra griega y la forma que adopta el cromosoma bacteriano en estados intermedios de replicación, no obstante pudiendo aparecer estructuras alternativas), que avanza en dirección a la región de ADN no duplicado dejando atrás los dos moldes de ADN de cadena simple donde se está produciendo la replicación.
Bidireccionalidad
El movimiento de la horquilla es bidireccional en la mayoría de los casos, es decir, a partir de un punto se sintetizan las dos cadenas en ambos sentidos. Esto ocurre en la mayoría de los organismos, pero se dan excepciones en algunos procariontes debido a que los mecanismos de replicación que tienen lugar dependen de la propia estructura de su material hereditario (si el ADN es circular, lineal, bicatenario o monocatenario). Así, en casos particulares como el ADN mitocondrial, algunos plásmidos y algunos genomas monocatenarios de fagos pequeños, la replicación se da unidireccionalmente pudiendo haber uno o dos orígenes de replicación.
No obstante, la replicación se puede considerar, de forma general, bidireccional.
La evidencia experimental del crecimiento bidireccional de la hebra de ADN viene dada por una técnica basada en el marcaje radiactivo del ADN usando timidina marcada con tritio. Primero se añade timidina marcada y luego sin marcar, y siguiendo el rastro de tritio se observa hacia dónde se ha replicado la molécula de ADN. También se puede, mediante el recuento de copias de genes marcadores, determinar si la replicación es unidireccional o bidireccional. Otras técnicas se basan en medir la distancia desde los ojos de replicación hasta los extremos de un ADN lineal (o circular convertido en lineal mediante enzimas de restricción).
Semidiscontinua
La replicación siempre se produce en sentido 5' → 3', siendo el extremo 3'-OH libre el punto a partir del cual se produce la elongación del ADN. Esto plantea un problema, y es que las cadenas tienen que crecer simultáneamente a pesar de que son antiparalelas, es decir, que cada cadena tiene el extremo 5' enfrentado con el extremo 3' de la otra cadena. Por ello, una de las cadenas debería ser sintetizada en dirección 3' → 5'.
Este problema lo resolvieron los científicos japoneses Reiji Okazaki y Tsuneko Okazaki en la década de 1960, al descubrir que una de las nuevas cadenas de ADN se sintetiza en forma de trozos cortos que, en su honor, se denominan fragmentos de Okazaki. Su longitud suele variar entre 1000 y 2000 nucleótidos en las bacterias y entre 100 y 400 nucleótidos en eucariontes.
La cadena que se sintetiza en el mismo sentido que avanza la horquilla de replicación se denomina hebra adelantada (en inglés, leading strand, que a veces se traduce por líder o conductora) y se sintetiza de forma continua por la ADN polimerasa, mientras que la que se sintetiza en sentido contrario al avance se denomina hebra rezagada o retrasada (en inglés, lagging strand), cuya síntesis se realiza de forma discontinua teniendo que esperar a que la horquilla de replicación avance para disponer de una cierta longitud de ADN molde.
La ADN polimerasa es la enzima que cataliza la síntesis de la nueva cadena de ADN a partir de desoxirribonucleótidos y de la molécula de ADN plantilla o molde que es la que será replicada. La enzima copia la cadena de nucleótidos de forma complementaria (A por T, C por G) para dar a cada célula hija una copia del ADN durante la replicación.
Modo de operación
En cada horquilla de replicación, la ADN polimerasa y otras enzimas sintetizan dos nuevas cadenas de ADN que son complementarias respecto a las 2 cadenas originales. Durante este proceso, la ADN polimerasa reconoce una base nucleotídica no apareada de la cadena original y la combina con un nucleótido libre que tiene la base complementaria correcta. Luego, la ADN polimerasa cataliza la formación de nuevos enlaces covalentes que ligan el fosfato del nucleótido libre entrante con el azúcar del nucleótido previamente agregado en la cadena hija en crecimiento. De esta forma, la ADN polimerasa sintetiza el esqueleto de azúcar-fosfato de la cadena hija.
En cada horquilla de replicación, la ADN polimerasa y otras enzimas sintetizan dos nuevas cadenas de ADN que son complementarias respecto a las 2 cadenas originales. Durante este proceso, la ADN polimerasa reconoce una base nucleotídica no apareada de la cadena original y la combina con un nucleótido libre que tiene la base complementaria correcta. Luego, la ADN polimerasa cataliza la formación de nuevos enlaces covalentes que ligan el fosfato del nucleótido libre entrante con el azúcar del nucleótido previamente agregado en la cadena hija en crecimiento. De esta forma, la
ADN polimerasa sintetiza el esqueleto de azúcar-fosfato de la cadena hija.
Las ADN polimerasas también realizan otras funciones durante el proceso de replicación. Además de participar en la elongación, desempeñan una función correctora y reparadora gracias a su actividad exonucleasa 3', que les confiere la capacidad de degradar el ADN partiendo de un extremo de éste. Es importante que existan estos mecanismos de corrección ya que de lo contrario los errores producidos durante la copia del ADN darían lugar a mutaciones.
Después de una serie de
experimentos con arvejas verdes y amarillas, observando como se transmitían las
características de los padres en varias generaciones, el botánico Gregor Mendel
planteó las leyes básicas de la transmisión de la herencia.
Durante sus observaciones Mendel
encontró que las características o rasgos almacenados de manera codificada en
los genes podían corresponder a características puras homocigotas o
características híbridas heterocigotas, en este último caso se trata de un par
de características alternativas de las cuales una es dominante (o sea que es la
que se manifiesta externamente en el organismo), y la otra es recesiva, o sea
que no se manifiesta externamente, pero permanece en la dotación genética y
puede hacerse visible en las siguientes generaciones.
Con
base en lo anterior Mendel formuló las siguientes leyes:
Si se
cruzan dos líneas puras (homocigotas) para un determinado carácter, los
descendientes de la primera generación son todos iguales entre sí (igual
fenotipo e igual genotipo) e iguales (en fenotipo) a uno de los progenitores.
Como cada uno de los progenitores es homocigoto, solo le puede pasar a la
descendencia el único alelo o variante del gen que porta.
Segunda
ley - Ley de la Segregación
Los
caracteres recesivos, al cruzar dos razas puras, quedan ocultos en la primera
generación (F1), reaparecen en la segunda (F2) en proporción de 1:3 uno a tres
respecto a los caracteres dominantes. Los individuos de la segunda generación
que resultan de los híbridos de la primera generación son diferentes
fenotipicamente unos de otros; esta variación se explica por la segregación de
los alelos responsables de estos caracteres, que en un primer momento se
encuentran juntos en el híbrido y que luego se separan entre los distintos
gametos
Ley de la
Dominancia.
Cuando se cruzan individuos que
difieren sólo en un carácter por ejemplo color de la semilla (dominante y
recesivo para este determinado carácter), la primera generación F1 será
semejante al progenitor que tiene el carácter dominante. En este caso se habla
de cruces monohíbridos
Ley de la transmisión
independiente o de la independencia de caracteres
Establece que los caracteres son
independientes y se combinan al azar. En la transmisión de dos o más caracteres,
cada par de alelos que controla un carácter se transmite de manera independiente
de cualquier otro par de alelos que controlen otro carácter en la segunda
generación, combinándose de todos los modos posibles.
Cuando se cruzan progenitores con
dos caracteres diferentes (ejemplo plantas puras es decir homocigotas con color
de las semillas amarillo dominante AA y verde recesivo aa y forma de la semilla
lisa dominante LL y rugosa recesiva ll), estos caracteres se trasmiten a la
descendencia en forma independiente. En este caso se habla de cruces dihíbridos.
Esto
se observa mejor mediante un cuadro de Punnet que permite visualizar las
posibles combinaciones para los cruces de caracteres.
Ejemplo: En los experimentos de
Mendel se encontraron:
plantas puras de arveja con semillas de color amarillo
dominante, o sea que sus alelos eran idénticos y se pueden denominar
convencionalmente AA
plantas puras de arveja con semillas de color verde recesivas,
las cuales denominaremos aa
plantas híbridas o
heterocigotas con semillas de color amarillo, Aa
plantas puras de arveja con semillas lisas como característica
dominante, LL
plantas puras de arveja con semillas rugosas como característica
recesiva, ll
plantas híbridas o
heterocigotas de arveja con semillas lisas, Ll
Aplicación de las leyes de Mendel en la
resolución de problemas sobre cruces monohibridos
Para
aplicar el cuadro de punnet analicemos primero el caso del cruce de plantas
homocigotas o puras de arveja con semillas amarillas dominantes AA y plantas
puras con semillas verdes recesivas aa (caso de cruce monohíbrido, o sea
aplicado a un solo carácter en este caso color de la semilla)
Se elabora una tabla o cuadro
con tres columnas y tres filas (cuadro de Punnet):
En
las celdas horizontales de color negro, van los alelos o genes aportados por el
padre (en este ejemplo el padre tiene un par de genes AA para el color de la
semilla) pero cada gameto solo recibe un gen para ese carácter por parte del
padre.
Entonces se coloca un gen A por
cada celda, o sea, un gen para la formación de cada gameto en el cruce.
Esto se explica de acuerdo con la
ley de la segregación Un par de genes es segregado (separado) en la formación de
los gametos.
En
las celdas verticales negras se colocan los alelos o genes que aportará la madre
a los gametos. De igual manera se cumple la ley de la segregación. Entonces en
cada celda se coloca un solo gen:
Las
celdas de color blanco corresponden a los gametos de los hijos que se formarán
en el cruce donde se restablecerá el número par de genes para cada gameto
Ejemplo: Si se cruzan semillas
homocigotas amarillas dominantes AA con semillas verdes homocigotas recesivas
aa, o sea que tenemos el caso AA x aa
En
las celdas blancas se formarán los gametos resultantes del cruce o sea la
combinación o entrecruzamiento de los genes aportados por el padre y la madre
para ese carácter (se combina el gen de la primera celda horizontal con el gen
de la primera celda vertical).
En este momento se restablece el
número par de genes en lo gametos formados (uno de cada progenitor)
El resultado
del cruce será: Genotipo: 100 % Heterocigoto Aa
Fenotipo: 100% Semilla de color amarillo.
(Ser puede explicar por la ley de la dominancia: un
gen del par determina la expresión fenotípica y enmascara al otro;
El
polen de la planta progenitora aporta a la descendencia un alelo o gen para el
color de la semilla, y el óvulo de la otra planta progenitora aporta el otro
alelo para el color de la semilla; de los dos alelos, solamente se manifiesta
aquél que es dominante (A), mientras que el recesivo (a) permanece
oculto.
Otro
ejemplo. Si se toman semillas heterocigotas lisas Ll y se cruzan con semillas
homocigotas rugosas ll.
Ll x
ll siguiendo el anterior procedimiento:
El
resultado del cruce será
Genotipo: 50 % Heterocigoto
Ll Fenotipo: 50% semilla de forma
lisa y 50% de semillas rugosas.
Interpretación de las leyes de Mendel - Ejemplos de
Cruces
Primera ley de Mendel o Ley
de la uniformidad de la primera generación filial (F1) o Ley de la Dominancia
Cuando se aparean o cruzan
organismos (fecundación) de raza pura (homocigotos) para un determinado carácter
, todos los individuos de la primera generación son iguales.
imagen
tomada de http://www.biotech.bioetica.org/ap1.htm
Ejemplo: Si se
cruzan arvejas amarillas AA con arvejas verdes aa toda la F1 resultante del
cruce será Aa de color amarillo. Aparece aquí el concepto de Dominancia y
Recesividad. Las arvejas
amarillas AA son dominantes sobre las arvejas verdes aa recesivas. La primera
generación o F1 es fenotípicamente amarilla y genotipícamente heterocigota Aa
imagen
tomada de http://www.biotech.bioetica.org/ap1.htm
Codominancia: La primera ley de Mendel se cumple
también para el caso en que un determinado gen de lugar a una herencia
intermedia y no dominante, como es el caso del color de las flores del "dondiego
de noche" (Mirabilis jalapa). Al cruzar las plantas de la variedad de flor
blanca con plantas de la variedad de flor roja, se obtienen plantas de flores
rosas. La interpretación es la misma que en el caso anterior, solamente varía la
manera de expresarse los distintos alelos
La segunda ley de Mendel también
llamada de la separación o segregación o disyunción de los
alelos
imagen
tomada de http://www.biotech.bioetica.org/ap1.htm
El experimento de
Mendel: Mendel tomó plantas procedentes de las semillas de la primera generación
del experimento anterior Aa y las polinizó entre sí. Del cruce Aa x Aa obtuvo
semillas amarillas y verdes en la proporción 3:1. Así pues, aunque el alelo que
determina la coloración verde de las semillas parecía haber desaparecido en la
primera generación filial, vuelve a manifestarse en esta segunada generación.
Interpretación del
experimento. Los dos alelos
distintos para el color de la semilla presentes en los individuos de la primera
generación filial, no se han mezclado ni han desaparecido , simplemente ocurría
que se manifestaba sólo uno de los dos. Cuando el individuo
de fenotipo amarillo y genotipo Aa, forme los gametos, se separan los alelos, de
tal forma que en cada gameto sólo habrá uno de los alelos y así puede explicarse
los resultados obtenidos
Retrocruzamiento
imagen
tomada de http://www.biotech.bioetica.org/ap1.htm
En
el caso de los genes que manifiestan herencia dominante, no existe ninguna
diferencia aparente entre los individuos heterocigóticos (Aa) y los
homocigóticos (AA), pues ambos individuos presentarían un fenotipo amarillo.
La prueba del retrocruzamiento, o
simplemente cruzamiento prueba, sirve para diferenciar el individuo homo del
heterocigótico. Consiste en cruzar el fenotipo dominante con la variedad
homocigota recesiva (aa). Si es homocigótico, toda la descendencia será igual,
en este caso se cumple la primera Ley de Mendel. Si es heterocigótico, en la
descendencia volverá a aparecer el carácter recesivo en una proporción del
50%
Tercera ley de Mendel o de la
herencia independiente de caracteres:
imagen
tomada de http://www.biotech.bioetica.org/ap1.htm
Hace referencia al caso de que se contemplen dos caracteres
distintos. Cada uno de ellos se transmite siguiendo las leyes anteriores con
independencia de la presencia del otro carácter.
El experimento de Mendel: Mendel
cruzó plantas de guisantes de semilla amarilla AA y lisa BB con plantas de
semilla verde aa y rugosa bb(Homocigóticas ambas para los dos
caracteres
Las
semillas obtenidas en este cruzamiento eran todas amarillas y lisas,
cumpliéndose así la primera ley para cada uno de los caracteres considerados , y
revelándonos también que los alelos dominantes para esos caracteres son los que
determinan el color amarillo y la forma lisa. Las plantas obtenidas y que
constituyen la F1 son dihíbridas (AaBb).
Segunda generación filial
F2
imagen
tomada de http://www.biotech.bioetica.org/ap1.htm
Se
cruzan entre sí plantas de la F1, teniendo en cuenta los gametos que formarán
cada una de las plantas
Los
alelos de los distintos genes se transmiten con independencia unos de otros, ya
que en la segunda generación filial F2 aparecen guisantes amarillos y rugosos y
otros que son verdes y lisos, combinaciones que no se habían dado ni en la
generación parental (P), ni en la filial primera (F1).
imagen tomada de
http://www.biotech.bioetica.org/ap1.htm
Interpretación del experimento: Los resultados de
los experimentos de la tercera ley refuerzan el concepto de que los genes son
independientes entre sí, que no se mezclan ni desaparecen generación trás
generación. Para esta interpretación fue providencial la elección de los
caracteres, pues estos resultados no se cumplen siempre, sino solamente en el
caso de que los dos caracteres a estudiar estén regulados por genes que se
encuentran en distintos cromosomas. No se cumple cuando los dos genes
considerados se encuentran en un mismo cromosoma, es el caso de los genes
ligados.
Hablar de herencia es hablar de genética, la ciencia que estudia la trasmisión hereditaria de los seres vivos a través de la reproducción. En otras palabras, herencia genética es la forma en que los progenitores dan a su descendencia los caracteres que ellos poseen.
Cualquier característica de un ser vivo que sea susceptible de ser trasmitida a su descendencia, la denominamos carácter hereditario.
El conjunto de características (no visibles) que un ser vivo hereda de sus progenitores se denomina genotipo, y aquellas que se hacen visibles en él se denomina fenotipo; este último viene determinado por el genotipo y las "condiciones ambientales" en las que el ser se ha desarrollado.
Forma de trasmisión de la herencia genética
La herencia genética es suministrada conjuntamente por los progenitores; es decir, el genotipo del nuevo ser está constituido por el aporte de ambos.
El macho aporta la herencia en loscromosomas del espermatozoide. La hembra aporta su herencia en los cromosomas del óvulo. La unión de espermatozoide y óvulo forma la célula huevo y dentro de ésta se encuentran los cromosomas de ambos; estos cromosomas son los que guardan la información de los caracteres hereditarios.
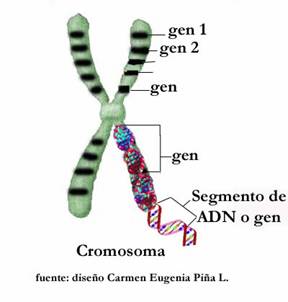
Los cromosomas son unos filamentos en los que se agrupan los genes, formados por secuencias de ADN y ARN.
Un gen es una unidad de trasmisión hereditaria que determinará, durante el desarrollo de un ser, la aparición o no de un determinado carácter. Así, pues, los elementos que determinan las características a heredar por un nuevo ser son los genes que están ubicados en los cromosomas de la célula huevo.
Los genes son pequeños segmentos de largas cadenas de ADN que determinan la herencia de una característica determinada, o de un grupo de ellas.
Los genes se encuentran localizados en los cromosomas en donde se disponen en línea a lo largo de ellos.
Cada gen ocupa en el cromosoma una posición, o “locus”.
En una célula, los cromosomas se agrupan por pares; en la célula huevo (origen de todas las que tendrá el nuevo ser), cada par posee un cromosoma de cada uno de los progenitores.
Los dos cromosomas que forman cada par son, generalmente, morfológica e intrínsecamente similares, y los genes situados en ellos en lugares homólogos, los llamados alelos o genes homólogos, son responsables de una determinada característica a la hora del desarrollo del nuevo ser.
Los genes alelos, entonces, son dos genes que ocupan el mismo lugar o “locus” en un par de cromosomas homólogos, es decir, en un par de cromosomas que tienen igual tamaño, forma y secuencia de genes.
También podemos decir que alelo es cada una de las variantes que puede presentar una secuencia de ADN polimórfica.
Cuando en alguna característica hereditaria interviene no solo un gen sino varios, que concurran en la trasmisión de este carácter, hablamos de alelismo múltiple.
Nuestra conocida arveja, los chícharos de Mendel.
Cuando los dos genes que determinan un carácter en sus respectivos cromosomas son idénticos, diremos que el carácter en cuestión se encuentra en homocigosis, el ejemplar es homocigoto; es decir, los genes aportados por sus progenitores son iguales.
Si por el contrario, ambos genes (para un mismo carácter) son diferentes, diremos que el ejemplar es heterocigoto, o, lo que es igual, el gen aportado por uno de los progenitores es diferente al aportado por el otro (insistimos, para un mismo carácter).
Si los dos genes que determinarán un carácter son distintos (heterocigoto) pueden ocurrir tres cosas en el momento de la formación del nuevo ser:
• 1. Que el carácter resultante lo sea según las directrices determinadas por uno de los genes.
• 2. Que el carácter resultante lo sea según las directrices del otro gen.
• 3. Que el carácter resulte una mezcla o superposición de ambos.
No todas las características se heredan de una manera tan simple como el color de las semillas de arvejas usadas por Mendel. Situaciones de herencia simple, como las analizadas por Mendel, en la que solamente interviene un par de genes, son las excepciones y no la regla.
Hoy sabemos que existen genes que se comportan respondiendo a la herencia mendeliana, mientras que otros quedan incluidos en la que llamaremos herenciano Mendeliana.
Entre estos últimos están el ligamiento, el crossing-over, la dominancia incompleta, los alelos múltiples, la codominancia y la herencia ligada al sexo y otras.
Interacciones génicas o genéticas
Luego de que los principios de la herencia fueron redescubiertos hacia el 1900, se realizaron diversos experimentos con el fin de probar su validez.
Dichos estudios comprobaron que los genes que se encontraban en el mismo “locus”, en un par de cromosomas homólogos (genes alelos) o en “locus” distintos (genes no alelos) podrían influir en la herencia de un rasgo. Estas interacciones se denominan interacciones génicas y pueden ser de los tipos: alélicas y no alélicas.
Los genes alelos; es decir, aquellos que se encuentran en el mismo “locus” en los cromosomas homólogos, pueden interactuar de diversas maneras y generar distintos mecanismos de herencia dominante, herencia recesiva, herencia intermedia, codominancia y series alélicas. Herencia dominante y herencia recesiva
Pelaje blanco, por un gen dominante.
Si prevalece un gen sobre el otro, diremos que el primero es dominante sobre el segundo, o que el segundo es recesivo respecto al primero. Diremos también que el ejemplar es portador delcarácter recesivo, pues lo posee aunque no lo manifiesta.
En la simbología genética, que usa letras para definir un carácter, las propiedades dominantes se escriben en mayúscula y las recesivas en minúscula.
El carácter dominante es siempre visible y oculta al recesivo.
El carácter recesivo puede permanecer latente durante generaciones y manifestarse cuando se den las condiciones de combinación adecuadas. Herencia intermedia
Como ya dijimos, dentro de los genes existen genes dominantes y genes recesivos. El gen dominante es aquel que se manifiesta y oculta al otro gen, que por este motivo se llama recesivo; lo que no quiere decir que no exista, sino que es mantenido en reserva oculto en la reproducción por el individuo.
Cuando los dos genes son semejantes se dice que es homocigótico. Significa que ese organismo tiene sus genes iguales para un mismo carácter. Cuando son diferentes se dice que es heterocigótico, es decir que sus genes para ese carácter (la altura, el color de pelo, color de ojos u otro) son distintos. Cuando ninguno de los genes es dominante se dice que se ha producido una herencia intermedia. Codominancia
Estado en que un gen expresa su característica en el heterocigoto de modo equivalente a su par. Los alelos del gen se expresan al mismo tiempo y de modo total en el heterocigoto. Dícese de los factores con la misma potencia hereditaria.
Este tipo de interacción se dilucidó estudiando la herencia de los grupos sanguíneos en el hombre.
En la especie humana se distinguen cuatro grupos sanguíneos: A, B, AB y O. Cuando uno de los progenitores es del grupo A y el otro del grupo B, el hijo puede ser del grupo AB, ya que los genes que determinan los grupos sanguíneos A y B se expresan de igual manera en el nuevo individuo, lo que se conoce como codominancia. Series alélicas
La mayoría de los genes alelos se pueden presentar en más de dos formas alternativas constituyendo las llamadas series alélicas. En ellas existen muchas variantes dentro del mismo par de genes, aunque un organismo diploide sólo puede presentar dos variantes de los genes que componen la serie alélica. Un ejemplo es el color del pelaje de los conejos, entre otros.
Las interacciones entre estos genes pueden darse en distintos niveles, distinguiéndose por ello fenómenos de epistasis, pleiotropía, genes modificadores y elementos genéticos transponibles. Epistasis
Es un tipo de interacción a nivel del producto de los genes no alelos. En una vía metabólica donde intervienen distintas enzimas, cada una de ellas transforma un sustrato en un producto, de manera que el compuesto final se obtiene por acción de varias enzimas. Cada una de estas está determinada por un gen, a lo menos.
Si uno de los genes que codificaba para alguna de las enzimas sufre una mutación y cambia, producirá una enzima defectuosa y el producto final no se obtendrá.
El efecto enmascarador sobre el fenotipo que tiene un gen sobre otro gen no alelo se denomina epistasis. En esta aparece un gen epistático y otro hipostático. El primero es el que enmascara el efecto del segundo.
Se distinguen distintos tipos de epistasis: dominante, recesiva, doble dominante y doble recesiva, y en cada una las proporciones clásicas se ven alteradas. Epistasis dominante. Se produce cuando el gen dominante es epistático sobre otro gen no alelo a él. Epistasis recesiva. En este tipo de interacción un gen recesivo actúa como gen epistático sobre otro gen no alelo. Epistasis doble dominante. En esta interacción, los genes presentes en los dos locus que intervienen en la característica, serán epistáticos en condición dominante.
Epistasis doble recesiva. Para que se produzca, los genes actúan como genes epistáticos deben estar en condiciones recesivas. Pleiotropía
Es un tipo de interacción entre genes no alelos que ocurre cuando la acción o cambio de un solo gen provocan la aparición de muchos fenotipos distintos. Genes modificadores
Son los que afectan la expresión de un gen diferente o no alelo. Un ejemplo de este tipo de interacción se observa en el color y distribución del manchado de los ratones. Elementos genéticos transponibles
Hasta 1960, se pensaba que los genes ubicados en los cromosomas eran estables e inmóviles, ya que se podían hacer mapas de su ubicación. Sin embargo, en 1947, B. Mc Clintock identificó, por primera vez, un grupo de genes que llamó elementos genéticos controladores.
En la actualidad se les denomina transponibles, elementos genéticos transponibles o genes saltarines. Uno de estos genes es un fragmento de ADN que puede moverse por todo el material hereditario de un organismo contenido en una célula. Este movimiento ocasiona cambios en el material hereditario que se traduce en la síntesis de distintos polipéptidos, lo que a su vez genera distintos fenotipos.
En algunas especies, los caracteres que se expresan como variaciones en cantidad o extensión, como el peso, la talla o el grado de pigmentación, suelen depender de muchos genes, así como de las influencias del medio.
Con frecuencia, los efectos de genes distintos parecen ser aditivos (se van sumando); es decir, parece que cada gen produce un pequeño incremento o descenso independiente de los otros genes.
Palmera enana.
Por ejemplo, la altura de una planta puede estar determinada por una serie de cuatro genes: A, B, C y D.
Supongamos que cuando su genotipo es aabbccdd, la planta alcanza una altura media de 25 cm, y que cada sustitución por un par de alelos dominantes aumenta la altura media en unos 10 centímetros.
En el caso de una planta que es AABBccdd su altura será de 45 cm, y en aquella que es AABBCCDD será de 65 centímetros.
En realidad, los resultados no suelen ser tan regulares. Genes diferentes pueden contribuir de forma distinta a la medida total, y ciertos genes pueden interactuar, de modo que la aportación de uno depende de la presencia de otro.
La herencia de características cuantitativas que dependen de varios genes se denomina herencia poligénica. La combinación de influencias genéticas y del medio se conoce como herencia multifactorial.
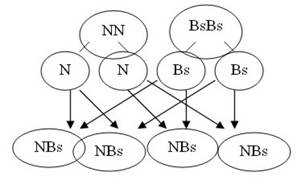
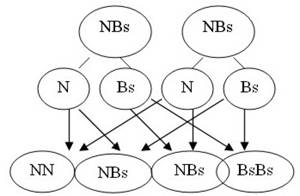
Corresponde a un tipo de herencia en que ambos genes son dominantes y se expresan simultáneamente (ejemplo: gallinas andaluzas).
Si se cruzan gallinas de plumas negras con gallinas de plumas blancas salpicadas de negro, en F1 (primera generación) se obtiene el ciento por ciento de gallinas con plumas negras y plumas blancas salpicadas de negro, simultáneamente, dando a las gallinas un color azulado (variedad andaluza).
Gallinas negras Genotipo: NN
Gallinas blancas salpicadas Genotipo BsBs
100 % gallinas andaluzas
Ahora, si se cruzan entre sí gallinas andaluzas, se obtiene:
25 % gallinas de plumas negras
50 % gallinas andaluzas
25 % gallinas de plumas blancas salpicadas, como lo demuestra el esquema siguiente.
Proporción fenotípica: 1 : 2 : 1 (25 por ciento negras, 50 por ciento andaluzas y 25 por ciento salpicadas).
Consanguinidad
Se habla de cruzamiento consanguíneo o endogamia cuando se cruzan entre si dos individuos con ascendentes comunes.
La consanguinidad, especialmente si es repetitiva, provoca taras y degeneración genética, ya que por consanguinidad tienen más probabilidades de aparecer en la descendencia genes letales recesivos. Es útil para fijar mutaciones e introducir nuevos caracteres que tienen lugar en caracteres recesivos.
¿Cuánto influye el genotipo en los deportistas?
La consanguinidad aumenta considerablemente la aparición de enfermedades recesivas y cuanto más próxima sea la relación familiar, mayor será el riesgo de que ambos miembros de la pareja hayan heredado el gen anormal del antepasado común.
La enfermedad autosómica recesiva más frecuente (1:2.500 recién nacidos vivos en poblaciones caucásicas) es la fibrosis quística, caracterizada por trastornos pancreáticos, respiratorios y de la sudoración, cuyo gene (CFTR) ha sido localizado en los brazos largos del cromosoma 7.
Herencia del sexo
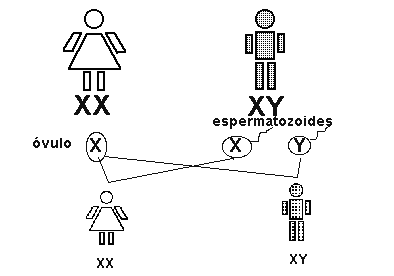
La especie humana posee 46 cromosomas, dispuestos en 23 pares. De ellos, solo un par es de cromosomas sexuales o heterocromosomas mientras que los 22 pares restantes son de cromosomas somáticos o autosomas.
Los cromosomas sexuales se han denominado X e Y. En los mamíferos, las células de los individuos machos contienen un par XY y las células de las hembras un par XX.
En la especie humana, como ya dijimos con una dotación de 46 cromosomas, cada célula somática femenina contiene 22 pares de autosomas más un par XX, y cada célula somática masculina contiene 22 pares de autosomas y un par XY.
La determinación sexual queda marcada en el momento de la fecundación y viene fijada por el tipo de gametos que se unen. Las mujeres sólo producirán un tipo de óvulo con 22 autosomas y un cromosoma sexual X, mientras que los varones formarán dos tipos de espermatozoides, el 50 por ciento serán portadores de un cromosoma X y el 50 por ciento serán portadores de un cromosoma Y.
Como la fecundacisn es producto del azar, un óvulo puede unirse a cualquiera de los tipos de espermatozoides, por lo que en la mitad de los casos se formarán hembras y en otro 50 por ciento se formarán machos.
Por lo tanto, queda claro que el cromosoma Y es el responsable de la masculinidad. Este cromosoma está casi vacío de genes, pero lleva suficiente información genética para el desarrollo sexual.
Herencia ligada al sexo
El cromosoma X es portador también de una serie de genes responsables de otros caracteres además de los que determinan el sexo.
Por eso decimos que la herencia de esos caracteres está ligada al sexo. Dos ejemplos bien conocidos son: el daltonismo y la hemofilia. Ver:Herencia ligada al sexo.
La simbología mendeliana
Mendel ideó una simbología que le permitió representar y entender los mecanismos que hacen posible la trasmisión de las características hereditarias de padres a hijos.
Los rasgos o caracteres estudiados por Mendel tenían siempre dos posibles manifestaciones fácilmente distinguibles, por ejemplo: el tamaño de la planta era alto o bajo, la textura de las semillas era lisa o rugosa, además, una de las alternativas de manifestación dominaba siempre a la otra.
También usaba dos letras para representar los "factores" que controla cada rasgo o característica estudiada.
En el tamaño de la planta, "A" representa el gen para tallo alto y "a" el gen que produce un caso enano. La característica dominante se denota siempre con letra mayúscula; la recesiva, con la misma letra pero minúscula.
Fenotipo: raza negra.
La genética actual, a partir del trabajo de Mendel, ha desarrollado algunos conceptos que son claves para entender los mecanismos de la herencia: fenotipo, genes alelos y genotipo. Fenotipo. Es la apariencia externa o visible de un organismo, todo lo que podemos observar y que es la expresión de la información genética. Por ejemplo, el color del cabello, de la piel, modelos de ojos, el tamaño y ubicación de las flores, la forma de las hojas y muchas otras. Genes alelos. Son segmentos específicos del ADN que determinan una característica hereditaria. Cada gen se ubica en uno de los cromosomas que forman el par homólogo, lo que permite su separación en diferentes gametos durante la meiosis.
En los estudios de Mendel los factores "A" y "a" son alelos por que ambos codifican para la misma característica (tamaño en la planta), aunque con expresiones distintas: alta y enana, respectivamente. Además, cada gen se ubica en un cromosoma de par homólogo, y están a la misma altura, en un lugar llamado “locus”. Genotipo. Es la constitución genética de un ser vivo, lo cual determina su fenotipo. El genotipo no es observable directamente, aunque sí se puede inferir a partir del análisis de las proporciones fenotípicas.
Cuando un organismo tiene genes alelos iguales, se dice que su genotipo es homocigoto (homo = igual). Existen dos tipos de homocigotos: dominantes y recesivos. El primero tiene sólo genes alelos dominantes (AA); el segundo lleva sólo genes alelos recesivos (aa).Cuando el individuo porta genes alelos distintos (Aa), se dice que su genotipo es heterocigoto.
El ciclo de Krebs (también llamado ciclo del ácido cítrico o ciclo de los ácidos tricarboxílicos) es una ruta metabólica, es decir, una sucesión de reacciones químicas, que forma parte de la respiración celular en todas las células aeróbicas. En células eucariotas se realiza en la mitocondria. En las procariotas, el ciclo de Krebs se realiza en el citoplasma, específicamente en el citosol.
En organismos aeróbicos, el ciclo de Krebs es parte de la vía catabólica que realiza la oxidación de glúcidos, ácidos grasos y aminoácidos hasta producir CO2, liberando energía en forma utilizable (poder reductor y GTP).
El metabolismo oxidativo de glúcidos, grasas y proteínas frecuentemente se divide en tres etapas, de las cuales, el ciclo de Krebs supone la segunda. En la primera etapa, los carbonos de estas macromoléculas dan lugar a moléculas de acetil-CoA de dos carbonos, e incluye las vías catabólicas de aminoácidos (p. ej. desaminación oxidativa), la beta oxidación de ácidos grasos y la glucólisis. La tercera etapa es la fosforilación oxidativa, en la cual el poder reductor (NADH y FADH2) generado se emplea para la síntesis de ATP según la teoría del acomplamiento quimiosmótico.
El ciclo de Krebs también proporciona precursores para muchas biomoléculas, como ciertos aminoácidos. Por ello se considera una vía anfibólica, es decir, catabólica y anabólica al mismo tiempo.
El Ciclo de Krebs fue descubierto el por el alemán Hans Adolf Krebs, quien obtuvo el Premio Nobel.